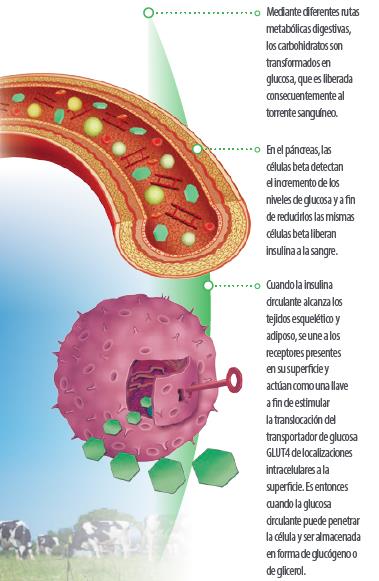
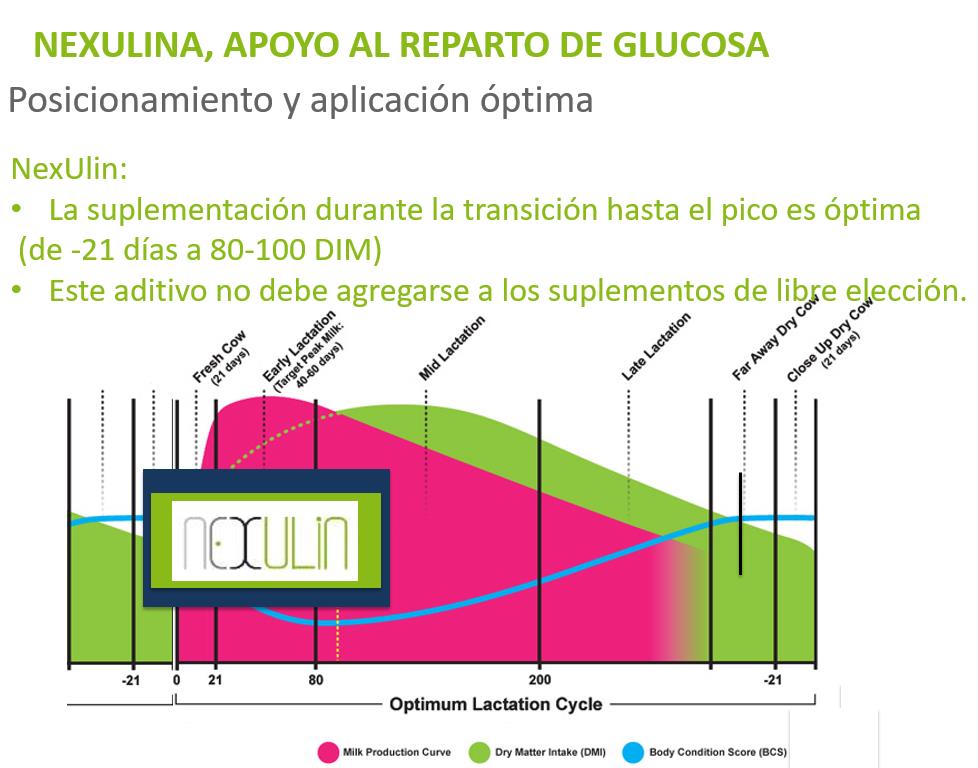
En vacas de leche, durante el período de transición entre gestación y lactación, la demanda masiva de glucosa por parte de la glándula mamaria hace que el reparto de glucosa sea un factor clave para mejorar los procesos productivos.
La insulina es la principal hormona involucrada en la regulación del mecanismo de glucosa y, como hormona anabólica que es, estimula la captación de glucosa por parte de las células, inhibiendo la gluconeogénesis hepática.
Investigaciones recientes han demostrado cómo determinados bioactivos pueden modular la liberación de insulina al flujo sanguíneo por parte del páncreas durante determinados períodos de desafío metabólico.
La regulación del metabolismo de la glucosa por la insulina
Los carbohidratos de la dieta aportan energía a los animales
.
Ahorro de glucosa – la Resistencia a la insulina en tejidos periféricos
Durante los estadios tempranos de la lactación, el lindar de respuesta a la insulina se ve incrementado y los tejidos esquelético y adiposo se vuelven más resistentes a la señal de esta hormona.
Así, la captación de glucosa por parte del tejido adiposo disminuye y existe más glucosa disponible para la glándula mamaria: este proceso recibe el nombre de ahorro de glucosa – glucose sparing – .
Inhibición de la gluconeogénesis por la insulina
El hígado produce glucosa mediante dos vías metabólicas: la gluconeogénesis y la glucogenólisis.
Cuando la gluconeogénesis se ve limitada y la demanda de energía es elevada, el hígado aporta Beta-hidroxibutirato (BHBA) mediante una oxidación de los NEFA (ácidos grasos no esterificados). El BHBA actúa entonces como fuente sustituta de energía en lugar de la glucosa.

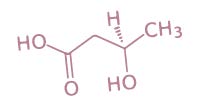
La Capsaicina disminuye la liberación de insulina
La oleorresina de Capsicum se extrae de los frutos de capsicum mediante hexano, siendo la capsaicina su principal ingrediente activo.
Su forma protegida a nivel ruminal es capaz de disminuir las concentraciones sanguíneas de insulina sin afectar a la concentración de glucosa (Oh et al., 2017).
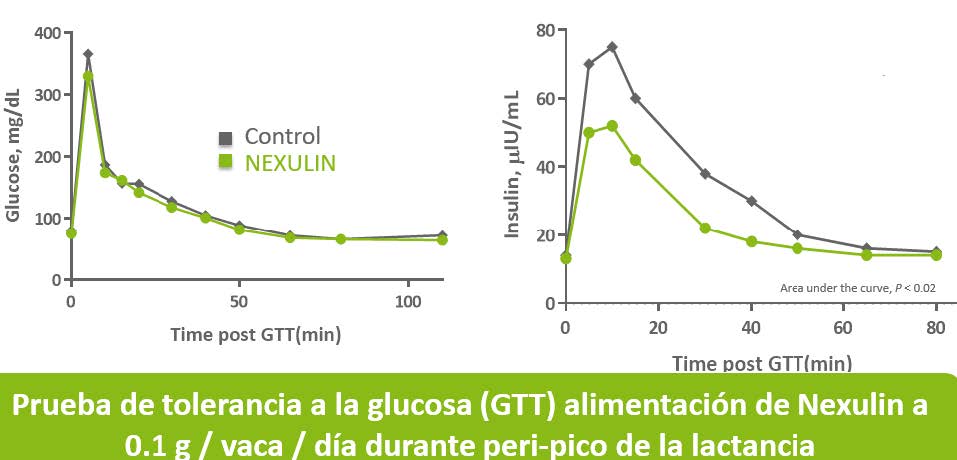
A su vez disminución de las concentraciones gracias a NEXULIN redirige la glucosa a la glándula mamaria y tejido de uterino generando un ahorro de glucosa que se perdía en tejidos periféricos.


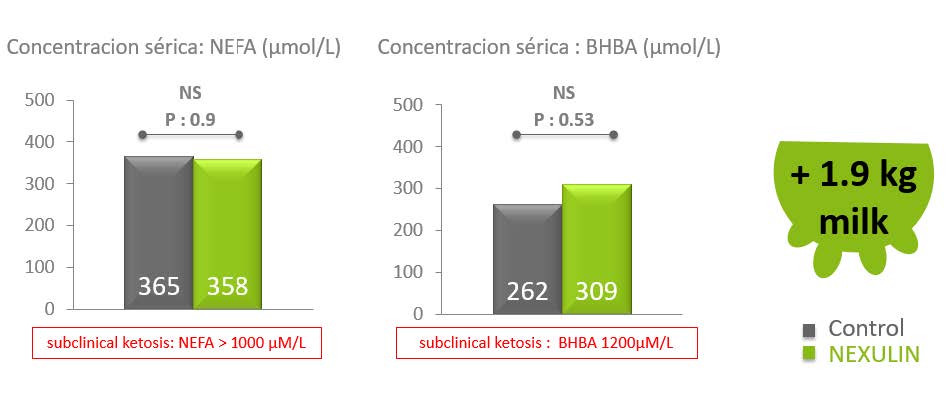
Así mismo se estudió que NEXULIN evita que en épocas de mayor demanda energética, donde existe la mayor posibilidad de originar Catabolismo energético o balance energético negativo, conllevando a la utilización de Grasa o triglicéridos para obtención de energía, y a su vez, aumenten en exceso los niveles de cuerpos cetónicos en sangre (NEFA o BHBA) originando cetosis o ketosis subclínica.
Se obtuvo que los niveles de cuerpos cetónicos en sangre son bajos como para producir cetosis subclinica
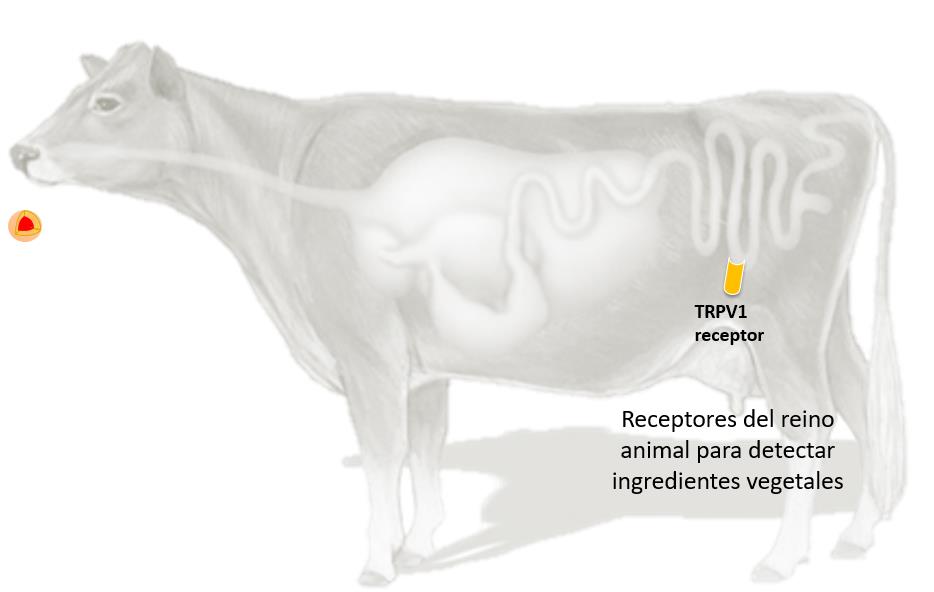
El efecto inhibitorio de la capsaicina parece relacionarse con la activación del receptor
TRPV1 (Transient Receptor Potential Caption Canel subfamilia V), también conocido como receptor de la capsaicina, cuya activación induce la secreción del péptido CGRP (Calcitonina Gene-Related).
Mediante la activación de TRPV1 en el intestino,el aditivo en base a capsicum by-pass* disminuye la liberación de insulina y, por tanto, evita la inhibición de la gluconeogénesis, apoyando en consecuencia el efecto de ahorro de Glucosa


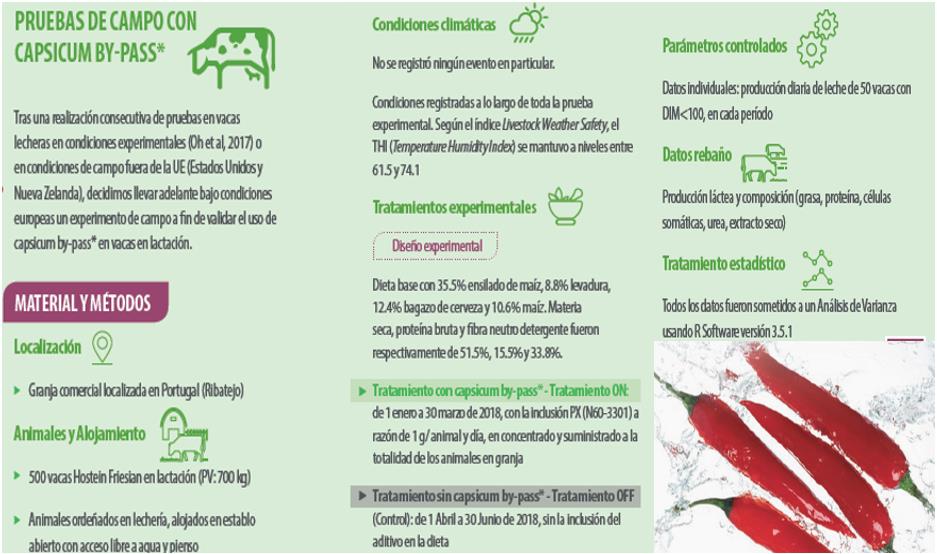
Resultados
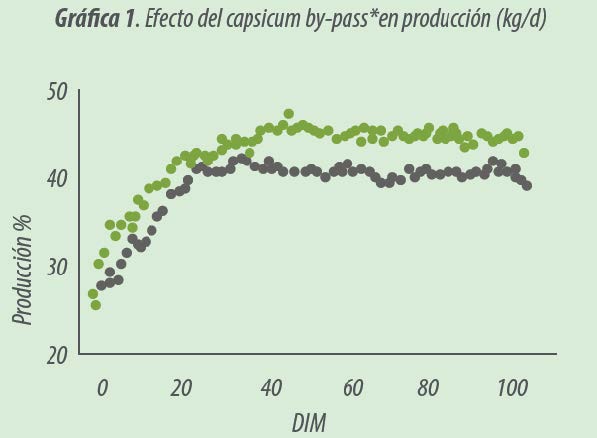
Las respuestas de producción a la inclusión de Capsicum by-pass* se presentan en la Gráfica 1.
Las vacas tratadas con capsicum by-pass* produjeron más leche (+3.55 kg/ d, P<0.001) que las vacas control.Este incremento en producción no conllevó un detrimento en la calidad de la leche.
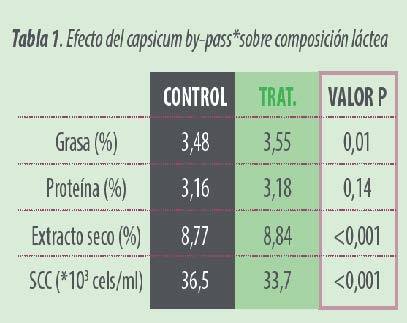
Las composiciones lácteas para ambos grupos se presentan en la Tabla 1.
El contenido en grasa incrementó significativamente (P<0.01) con la suplementación de capsicum by-pass* ,manteniéndose el contenido proteico.
El recuento de células somáticas fue significativamente (P<0.001) inferior con capsicum by-pass. A pesar de los ya excelentes resultados obtenidos en el grupo control.
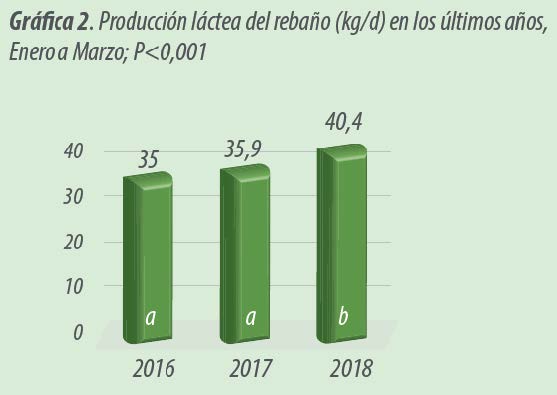
Al estudiar los datos históricos de la granja, para un período similar (enero a marzo), existe un incremento significativo en la producción diaria de leche por parte del rebaño (P<0.001).
Durante el año de la prueba experimental se incrementó la producción en 4.63 kg/ d con respecto a 2017 y en 5.33 kg/ d en comparación a 2016 (Gráfica 2).
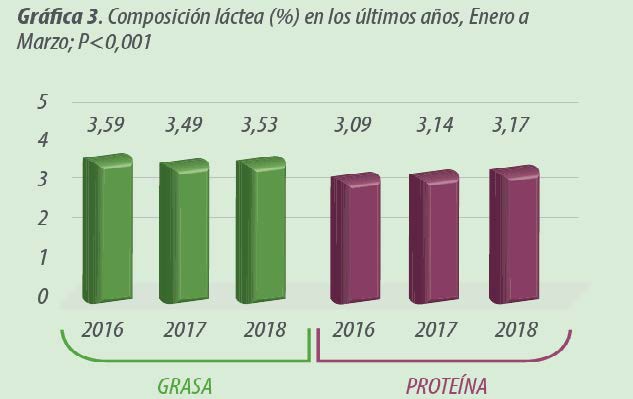
En lo que respecta a calidad de leche, la proteína láctea (en %) fue significativamente superior en el año delexperimento (Gráfica 3) mientras que no se observaron diferencias significativas en el porcentaje graso.